在肺炎双球菌的转化实验中, 能使R型细菌转化为S型细菌的转化因子是
| A.R型细菌的蛋白质 | B.S型细菌的蛋白质 |
| C.S型细菌的 DNA | D.R型细菌的糖类 |
肺炎双球菌的转化实验和噬菌体侵染细菌的实验,证明了()
| A.DNA是主要的遗传物质 |
| B.蛋白质是遗传物质 |
| C.RNA是遗传物质 |
| D.DNA是遗传物质 |
图2为用标记的T2噬菌体侵染大肠杆菌(T2噬菌体专性寄生在大肠杆菌细胞内)的实验,据图判断下列叙述正确的是( )
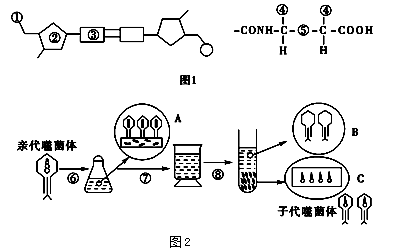
| A.图2中用35S、32P分别标记噬菌体的蛋白质、DNA,标记元素所在部位分别对应于图1中的⑤① |
| B.锥形瓶中的培养液是用来培养32P标记的T2噬菌体的,其营养成分中应含有32P |
| C.图2中的⑥⑦分别代表搅拌、离心过程,其中⑦的目的之一是让B上清液中析出质量较轻的T2噬菌体颗粒 |
| D.噬菌体增殖需要细菌提供模板、原料和酶等 |
艾弗里在格里菲思实验的基础上进一步实验探究“转化因子”,A、B、C、D为四个培养有R型细菌的试管,艾弗里将从S型活细菌中提纯得到的DNA、蛋白质、多糖分别加入试管中,还有一组是用DNA酶分解S型活细菌中提取的DNA之后将产物加入试管中,实验过程如图所示。经过培养后进行检查,发现没有S型细菌形成的试管是
用噬菌体侵染体内含32P的细菌,细菌解体后,含32P的应是( )
| A.子代噬菌体的DNA | B.子代噬菌体的蛋白质外壳 |
| C.子代噬菌体的所有部分 | D.以上都不正确 |
注射后能使小白鼠因患败血病而死亡的是 ( )
| A.R型肺炎双球菌 |
| B.加热杀死后的R型肺炎双球菌 |
| C.加热杀死后的S型肺炎双球菌 |
| D.加热杀死后的S型肺炎双球菌与R型细菌混合 |
在肺炎双球菌的转化实验中, 能使R型细菌转化为S型细菌的转化因子是
| A.R型细菌的蛋白质 | B.S型细菌的蛋白质 |
| C.S型细菌的 DNA | D.R型细菌的糖类 |
肺炎双球菌的转化实验和噬菌体侵染细菌的实验,证明了()
| A.DNA是主要的遗传物质 |
| B.蛋白质是遗传物质 |
| C.RNA是遗传物质 |
| D.DNA是遗传物质 |
某科研小组对“噬菌体侵染细菌实验”过程中搅拌时间与放射性强弱关系进行了研究,结果如下图所示。下列分析不正确的是
| A.实验过程中被侵染的细菌基本未发生裂解 |
| B.实验过程中充分搅拌能使所有的噬菌体与细菌脱离 |
| C.实验结果表明,做“噬菌体侵染细菌实验”的适宜搅拌时间为2 min左右 |
| D.若搅拌4 min时被侵染的细菌下降为90%,则上清液中32P的放射性会增强 |
某同学分离纯化了甲、乙两种噬菌体的蛋白质和DNA,重新组合为“杂合”噬菌体,然后分别感染大肠杆菌,并对子代噬菌体的表现型作出预测,见表。其中预测正确的是( )
| “杂合”噬菌 |
实验预期结果 |
|
| 预期结果序号 |
子代表现型 |
|
| 甲的DNA+ 乙的蛋白质 |
1 |
与甲种一致 |
| 2 |
与乙种一致 |
|
| 乙的DNA+ 甲的蛋白质 |
3 |
与甲种一致 |
| 4 |
与乙种一致 |
A.1、3 B.1、4 C.2、3 D.2、4
在“噬菌体侵染细菌”的实验中,如果对35S标记的噬菌体实验组(甲组)不进行搅拌、32P标记的噬菌体实验组(乙组)保温时间过长,则会出现的异常结果是( )
| A.甲组沉淀物中也会出现较强放射性,乙组上清液中也会出现较强放射性 |
| B.甲组上清液中也会出现较强放射性,乙组上清液中也会出现较强放射性 |
| C.甲组沉淀物中也会出现较强放射性,乙组沉淀物中也会出现较强放射性 |
| D.甲组上清液中也会出现较强放射性,乙组沉淀物中也会出现较强放射性 |
1952年赫尔希和蔡斯用35S和32P分别标记T2噬菌体时,做法是 ( )
| A.分别用35S和32P的人工培养基培养T2噬菌体 |
| B.分别将35S和32P注入鸡胚,再用T2噬菌体感染鸡胚 |
| C.分别用35S和32P的培养基培养细菌,再分别用上述细菌培养T2噬菌体 |
| D.分别用35S和32P的动物血清培养T2噬菌体 |
肺炎双球菌的转化实验和噬菌体侵染细菌的实验,证明了( )
| A.DNA是主要的遗传物质 | B.蛋白质是遗传物质 |
| C.RNA是遗传物质 | D.DNA是遗传物质 |
将加热杀死的S型细菌与R型活细菌相混合后,注射到小鼠体内,在小鼠体内S型和R型细菌含量变化情况如图所示。下列有关叙述中错误的是 ( )

| A.在死亡的小鼠体内可分离出S型和R型两种活细菌 |
| B.曲线ab段下降是因为部分R型细菌被小鼠的免疫系统所消灭 |
| C.曲线bc段上升,与S型细菌使小鼠发病后免疫力降低有关 |
| D.S型细菌数量从0开始增多是由于R型细菌基因突变的结果 |
 粤公网安备 44130202000953号
粤公网安备 44130202000953号